Long strands of collagen molecules are the most abundant protein in our bodies. They have a unified presence everywhere within us that is analogous to every water molecule on our planet: There is one continuous body of water with no boundaries. Fog, raindrops, puddles, rivulets, streams, lakes, rivers, and all of the variously named oceans are one unified, endlessly circulating collective of water, and it penetrates every nook and cranny on the globe.
And so it is with our collective distribution of collagen: Fascia, tendons, ligaments, the loose network under our skin, down to the microscopic filaments that pass through cell walls to unite with every cell’s cytoskeleton. Its consistencies vary from compact and dense to thin and light to highly flexible and spongy structures, and like water it is simply everywhere.
Unfortunately, our typical anatomical textbooks are very limiting and misleading in their depictions of connective tissue structures and their continuity. We are given illustrations of red muscle structures with white connective tissue borders and tendons. Or we see drawings of skeletal structures with ligament connections that appear to be fixed to bones across joints rather like pieces of tape. This pictoral sense of things is false, and it can lead us to therapeutic strategies that may not be in keeping with the way things are actually put together and the functional relationships that they form.
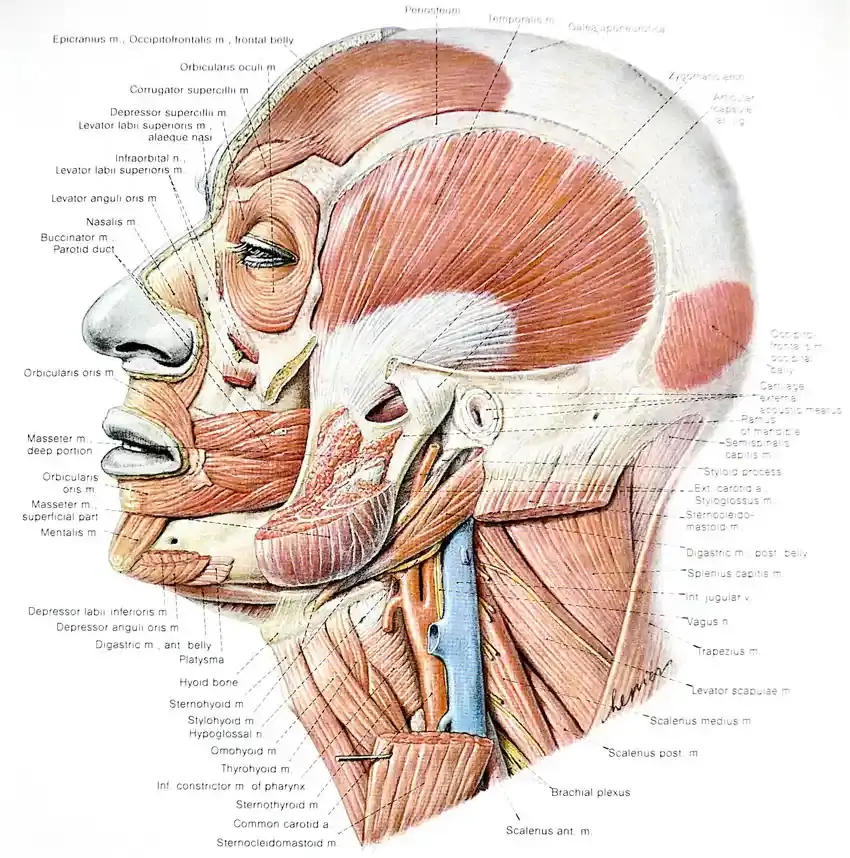
Sobota, Atlas of Human Anatomy; copyright Urban and Schwartzenberg, 1983
This is a typical textbook illustration. Note how compartmentalized and tidy muscles and connective tissue structures appear. Most of the interconnectedness is absent.
Let us do some tracing, beginning with the bones. A “bone” is nothing like the skeletons we see in our classrooms, which are dried out and with all connective tissue removed. In truth a bone is a connective tissue mold into which are deposited the mineral salts—primarily calcium—that give it a springy solidity. The outer layer of this mold is the periosteum that encases the bone. This layer is then continuous with the ligaments that connect each bone to the next bone across a joint; ligaments are just thickenings of the periosteum, which transition into the periosteum of the next bone.
Then let us look at tendons, which in our illustrations seem to simply appear at the ends of muscle bellies and are “attached” to a bone’s surface like they were glued down to a specific location. Well, no, not really. Every single muscle cell is encased in its own long tubular sheath within the muscle belly. The seemingly separate tendons are where these tubes pack together at the end of the muscle cell’s structure; and they are not “glued down” to the bone, but are infused into that bone’s periosteum, which continues on to further ligament thickenings and more tendons emerging from more muscles. And so on, from bone to bone to tendon to muscle belly throughout the entire skeletal-muscular continuum.
We find the same picture emerging when we consider all other tissue structures in our bodies. Every cell in a liver, for instance, is surrounded by its own tiny sac of connective tissue which is in turn connected to all surrounding liver cell sacs, which ultimately are all thus connected to the larger, thicker sac that surrounds the liver as a whole. Then this larger liver sac is fused into sheets and folds of connective tissue—the peritoneum—that are also suspending the intestines and glands within the abdomen. This suspension is what keeps all the internal organs in their functional place, and not falling in a heap into the pelvic bowl.
Indeed it is the same with all other structural systems in the body. Every blood vessel is a tube of muscle and collagen, every lymph vessel is a collagen tube, every nerve bundle, and separate axon is sheathed in their tubing, and all of these hundreds of thousands of miles of tubing are anchored in connective tissue meshes that keep them in their proper places.

Carla Stecco, Functional Atlas of the Human Fascial System; copyright Elsevier Ltd., published 2015.
This is a real neck in dissection. Note the extensive interconnections of all structures, with connective tissue matrix uniting all layers of tissue.
Next, let’s look at the elaborate mechanical transfer of forces that result in every one of our movements. Functionally there is simply no way to separate the bracing, shortening, and lengthening activities of our muscles from the meshes of connective tissue that surround them and fuse them into the bones. All of the connective tissues I have been describing are ultimately unified into a single matrix, a three-dimensional spider web, and every lengthening or shortening of any group of muscle fibers anywhere creates a shift in tension lines throughout the entire web.
This is why it cannot be said (despite the testimony of many kinesiology textbooks) that any one muscle ever does any one thing, or that any muscle can ever act alone. All blends of lengthening and shortening of muscle fibers result in a shift of mechanical tensions within the whole webbing; there is a shape-changing that is then transferred to the skeleton as a whole, which produces the complex combinations of secure bracings and movements of our bodies in the field of gravity. The muscular shifts from bracings to shortenings to lengthening are global and kaleidoscopic, shifting in milliseconds from one configuration to the next. Muscle fibers and connective tissue structures are all working in perfect harmonies of space and time to get us from one position to the next in our movings about.
There is yet another connectedness that this tissue confers upon us. Collagen fibers are not only structural—they have a dynamic energetic property as well. The entire webbing is a semiconducting material that absorbs electromagnetic fields. Moreover, they are of a particular sort of semiconductors that are referred to as piezo-electric (piezo=self-generating). Crystals, for instance, are piezo-electric: when you apply pressure to their matrix they generate current. And they are very efficient conductors of this current as well; this is what makes them so useful in electronic inventions. Likewise, when collagen structures are compressed, stretched, distorted they generate their own electrical field which the also conduct throughout their entire matrix. So this means that our collective of cells is not only held together physically with the woven textile of connective tissue, but are all also bathed in the electromagnetic current it is generating and conducting.
It is a curious fact that our bodies appear to be producing more metabolic energy than can be accounted for by strictly measuring the calories of our food intake. Perhaps it is this self-generated current of the connective tissue matrix that accounts for the discrepancy.
This finally leads us to a last, far more wide-reaching connectedness we are given by the remarkable nature of our connective tissue matrix. Our whole organism is generating an electromagnetic field that surrounds us and extends beyond our physical boundary. We are, in effect, magnets. This field can be measured with sufficiently sensitive instruments; perhaps being gifted with a little wider bandwidth of visual wavelength sensitivity is what allows some people to clearly see the auras in this field. And of course when we gather together in proximity we are merging fields with one another. I suspect this is what we called “vibes” back in the day,
And of course, each of our individual magnets is embedded within the far larger magnetic field of the Earth, which is visibly measurable with a compass. And in turn the Earth’s field is embedded in the solar magnetic field of the sun, which is embedded within our galactic field, which ultimately merges into the vast fields of the cosmos of which we are each a humble part.
Connected indeed.
Copyright January 10, 2024, by Deane Juhan